| |
introduction
migration
matrix
zebrafish
|
|
Drosophila egg chamber development as a model for organ morphogenesis
During development, discrete organs and entire body plans emerge from the coordinate actions of
individual cells. These complex morphogenetic events require dynamic regulation of cell shape,
polarity, and adhesion across cell populations. Our lab seeks to understand how these diverse
cellular behaviors are orchestrated to produce an organ's functional shape.
|
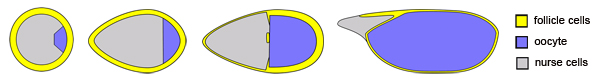 - Changes in egg chamber morphology over time (images not drawn to scale)
|
|
To this end, we are using the Drosophila egg chamber as a highly tractable system to investigate
the cellular control of organ morphogenesis. Egg chambers are multi-cellular structures within fly
ovaries that consist of an inner germ cell cluster surrounded by an epithelial layer of follicle
cells. This simple organ's function is to nurture and pattern the oocyte growing within, as each
egg chamber will give rise to a single egg. Despite the fact that it is an adult structure, the egg
chamber undergoes complex morphogenetic changes that rival those seen in embryos. We are
currently investigating how the egg chamber is transformed from a sphere to an ellipsoid as it
grows. As is detailed in the subsequent pages, we are particularly interested in the role that
collective cell migration and basement membrane remodeling play in this process.
|
